【协和医学杂志】感染在自身免疫性疾病中的作用机制
2023-10-17 协和医学杂志 协和医学杂志 发表于上海

本文通过回顾最新文献,探讨病原微生物及相关免疫通路可能促进自身免疫发展的机制,以及这些感染因子触发或加重AID的相关研究进展,以期加深临床对感染因子与AID发病潜在关联的认识。
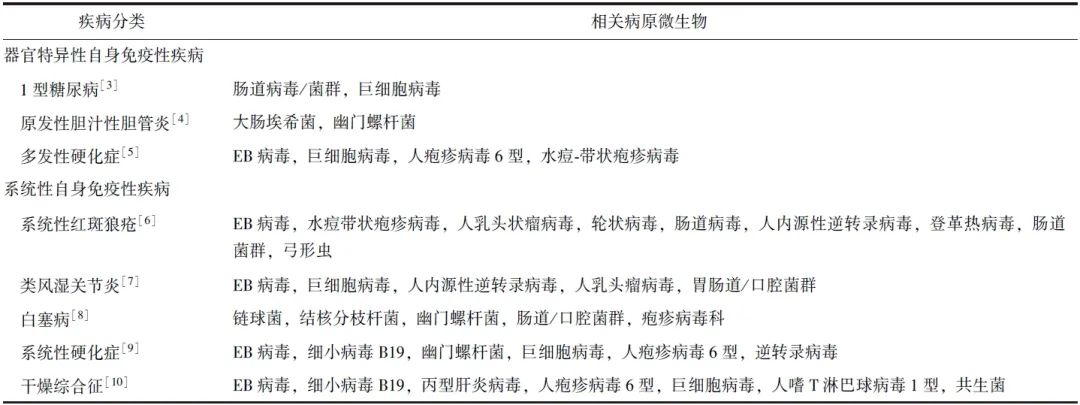
自身免疫性疾病(AID)可分为器官特异性和系统性两大类,其病因和发病机制尚未完全阐明。在过去几十年中,关于病原微生物与自身免疫/AID之间关系的研究已广泛开展(表1)。
表1 自身免疫性疾病及相关病原微生物
越来越多的证据表明微生物感染是人类AID的潜在触发因素,尤其是在具有遗传倾向的人群中[1],而COVID-19大流行使得感染与人类自身免疫之间的关联表现得更为强烈。
病原微生物可通过不同机制参与自身免疫的发生和进展,包括分子模拟、旁观者激活和表位扩展等[2]。本文通过回顾最新文献,探讨病原微生物及相关免疫通路可能促进自身免疫发展的机制,以及这些感染因子触发或加重AID的相关研究进展,以期加深临床对感染因子与AID发病潜在关联的认识,为进一步了解AID发病机制提供线索,以助于开发有效的预防性治疗策略,最大限度降低AID的发病风险。
1 发病机制
1.1 分子模拟
分子模拟是感染或化学因素诱导自身免疫的主要机制之一,即外源肽和自身肽之间存在相似性,导致易感个体的外源抗原激活自身反应性T/B细胞[11]。
越来越多的证据表明,病原微生物感染可通过分子模拟和交叉反应促进自身免疫和疾病的发生、发展。如患者感染A组链球菌后,可产生针对心脏、关节、大脑和皮肤组织蛋白反应的抗体,进而介导包括急性风湿热在内的后遗症[12]。
此外,交叉反应也可发生于T细胞水平。微生物抗原与自身抗原具有抗原相似性,抗原提呈细胞可将这些自身抗原呈递给交叉反应性T细胞,进而发生自身免疫反应。
1.2 旁观者效应
在适应性免疫反应中,T细胞可对同源抗原产生特定的抗原免疫反应。CD8+ T细胞激活依赖于T细胞受体(TCR)介导的刺激,通过与主要组织相容性复合体(MHC)Ⅰ类呈递的同源肽结合。
然而,在具有遗传易感性的个体中,微生物感染可触发炎症信号,若无同源抗原的刺激,细胞因子等非特异性因素,如白细胞介素(IL)-15和IL-18也可激活记忆T细胞,即旁观者T细胞激活,并进一步分化为效应细胞,介导免疫病理损伤导致组织破坏[13]。此外,记忆CD4+ T细胞也可通过多种细胞因子的TCR非依赖性刺激进行旁观者激活[13]。
1.3 表位扩展
在许多免疫反应中,适应性免疫系统识别的初始抗原和抗原表位是有限的。在慢性自身免疫或炎症反应中,会产生针对其他表位甚至其他抗原分子的免疫反应,此过程被称为表位扩展。
表位扩展可同时发生于T/B细胞水平、单个抗原上的多个表位之间或不同抗原分子的表位之间。内吞作用、抗原呈递和体细胞超突变是推动表位扩展和扩大AID免疫反应的重要分子机制[14]。表位扩展是免疫系统所识别表位的多样化,有助于提高适应性免疫效率,但同时也会导致自身免疫反应的发生。
1.4 其他
病原体可对受感染细胞造成直接损害,导致感染细胞坏死。发生微生物感染时,自身抗原可被抗原呈递细胞摄取和加工,在此过程中,炎症信号导致抗原呈递细胞对自身抗原的处理方式发生改变,暴露出隐藏的表位,隐匿抗原呈递给自身反应性T细胞,进而启动自身免疫反应[2]。此外,病原体衍生的超抗原,如葡萄球菌肠毒素,也可直接将MHC分子与TCR交联,无需特定的肽即可激活T细胞[15]。
2 感染与器官特异性自身免疫性疾病
2.1 1型糖尿病
1型糖尿病(T1DM)多见于儿童和青少年,其原因是产生胰岛素的胰岛β细胞遭到破坏,导致胰岛素产生不足。截至目前,与T1DM相关的最有力证据指向肠道病毒感染(表1),尤其是柯萨奇病毒,其是最易从肠道黏膜传播至胰腺的肠道病毒之一[16]。研究表明,柯萨奇病毒B感染的直接细胞病变效应是导致β细胞死亡的主要原因[17]。
此外,柯萨奇病毒B在胰腺内的持续存在可促进固有免疫激活及慢性炎症,且预先存在的自身反应性细胞毒性T细胞可对β细胞产生进行性自身免疫性破坏[16]。分子模拟在T1DM患者发病过程中也发挥重要作用,如与胰岛素类似的梭状芽胞杆菌肽可与胰岛素受体类似的柯萨奇病毒肽结合,被T1DM患者的T细胞受体序列识别[18],进而启动胰岛自身免疫。
2.2 原发性胆汁性胆管炎
原发性胆汁性胆管炎(PBC)是一种慢性胆汁淤积性肝病,以肝内进行性的小胆管炎症、免疫损伤和肝纤维化为特点。流行病学证据表明,细菌感染与PBC有关,其中尿路感染增加了PBC的发病风险,而大肠埃希菌是尿路感染患者中最常分离出的病原体[19]。
大肠埃希菌丙酮酸脱氢酶复合体E2亚单位(PDC-E2)与人类PDC-E2的分子结构高度相似,可通过分子拟态和交叉反应等机制打破人体对自身PDC-E2抗原的免疫耐受,产生抗线粒体抗体,导致PBC的发生[20]。
此外,人类和大肠埃希菌PDC-E2的共有序列也可被T细胞识别,在促进PBC发病机制中发挥不可忽视的作用。然而,由于PDC-E2在系统遗传学方面从原核生物到高等生物均高度保守,其他细菌通过分子模拟参与PBC发病的可能性尚不能排除[20]。
2.3 多发性硬化症
多发性硬化症(MS)是一种起源于中枢神经系统的AID,炎症、脱髓鞘和轴突变性是引起MS临床表现的主要病理学机制。目前MS的病因学研究大多集中于EB病毒。研究已表明EB病毒感染与MS发病风险增加具有直接因果关系[21]。
分子模拟在MS的发病机制中具有重要作用。MS患者血清中存在针对EB病毒核抗原1(EBNA1)的抗体,与中枢神经系统蛋白神经胶质细胞黏附分子、热休克蛋白、α-晶状体球蛋白、髓鞘碱性蛋白和氯离子通道蛋白等人体蛋白存在交叉反应,且这些连续的交叉反应抗原表位可能与表位扩展有关[9,22]。
然而,EB病毒可能在MS的不同阶段发挥不同作用,从发病时通过分子模拟诱导自身反应到疾病后期增强外周B细胞活化和呈递,以及靶组织的中枢神经系统免疫反应[23]。开发针对EB病毒的疫苗可能是未来治疗和预防MS的新契机[24]。
3 感染与系统性自身免疫性疾病
3.1 系统性红斑狼疮
系统性红斑狼疮(SLE)是一种病因不明的几乎可影响所有器官的慢性AID,各种传染性病原体尤其是病毒与SLE存在相关。近50%的SLE风险等位基因可被EB病毒转录因子EBNA2蛋白靶向结合,包括自身免疫相关位点,表明EB病毒可干扰免疫反应[25]。
另一方面,机体对EB病毒蛋白的异常反应促进了核蛋白交叉反应自身抗体的表达和表位扩展的发生,导致抗体进一步结合Sm和Ro60蛋白上的其他表位以及其他核抗原[26]。单纯疱疹病毒(HSV)和巨细胞病毒(CMV)等人类疱疹病毒也与SLE存在相关[27]。
尽管HSV与SLE的发病相关性不如其他疱疹病毒特别是EB病毒显著,但由于中枢系统感染死亡率高,且与神经精神狼疮鉴别诊断难度大,故HSV在SLE中的作用不应被低估。此外,当大剂量免疫抑制剂治疗神经精神狼疮时,更易合并严重HSV感染[28]。最新研究也揭示,SLE患者皮肤微生物群和产道微生物群的组成和丰度变化与SLE的发病机制存在相关[29]。然而,相较而言,病毒可能在SLE发病中的作用更加突出。
3.2 类风湿关节炎
类风湿关节炎(RA)是一种常见的AID,以滑膜炎症和关节损伤为主要临床表现。微生物组在RA的发生发展中起重要作用,但其具体机制尚未明确,目前研究主要涉及胃肠道的普雷沃氏菌和口腔的牙龈卟啉单胞菌[30]。
牙龈卟啉单胞菌通过介导细菌和宿主蛋白瓜氨酸化诱发RA的自身免疫机制,普雷沃氏菌copri可能通过辅助性T细胞17(Th-17)激活免疫反应并引起RA免疫紊乱[31-32]。
普雷沃氏菌copri的定植伴高纤维饮食促进琥珀酸盐的过量产生,可通过促进巨噬细胞炎症反应导致RA的发生发展[33]。然而,不同研究在探究肠道微生物组与RA因果关系得出的结果似乎存在争议,关于普雷沃氏菌等微生物组在RA中发挥的作用有待进一步研究确认。
此外,变形杆菌、EB病毒及结核分枝杆菌等致病菌可能通过介导分子模拟促进RA的发生发展[7]。
3.3 白塞病
白塞病(BD)是一种异质性的慢性血管炎,患者可出现口腔溃疡、生殖器溃疡、葡萄膜炎和皮肤损害等多组织/器官症状。研究表明,病原微生物在BD发生发展中起关键作用[8],其中疱疹病毒科成员是研究最多的病毒因子,尤其是HSV[8]。
HSV诱导的BD样小鼠模型与人类最相似,这些症状可能发生于免疫反应的基础上,而非病毒感染[8]。其中HSV被膜蛋白VP16与人类热休克同源71 kDa蛋白(Hsc71)存在IgA和IgG交叉反应[34]。Hsc71对维持细胞稳态[35]以及向CD4+和CD8+ T细胞呈递抗原至关重要[35-36]。
人体大量暴露于HSV组分,可引起自身免疫反应和人Hsc71活性受损,导致BD自身反应性免疫细胞的活化和炎症反应加重。血链球菌GroEL蛋白和人重组核异质性核糖核蛋白(hnRNP)的8个同源表位可与血清IgA发生交叉反应,进而启动和维持自身免疫[37]。
BD患者对链球菌抗原刺激可表现为迟发性皮肤反应和炎症反应,基于此开发的用于BD诊断的23价肺炎球菌多糖疫苗抗原刺激法[38]也表明,病原微生物在BD发生发展中发挥重要作用,该法具备较高的临床转化价值。
3.4 系统性硬化病
系统性硬化病(SSc),又称硬皮病,以皮肤等多组织/器官纤维化为主要病理表现。研究报道,EB病毒、逆转录病毒、CMV、细小病毒和幽门螺杆菌等病原体感染可能均与SSc发病有关[39]。其中,逆转录病毒蛋白与SSc特异性自身抗原U1核糖核蛋白及Scl-70具有同源序列;人免疫缺陷病毒1型的保守pol内源序列与SSc患者抗U1核糖核蛋白抗体阳性及皮肤受累程度相关。
CMV与人疱疹病毒6型可能共同诱导纤维化和凋亡相关因子的表达改变,在SSc中或发挥联合作用[40]。人类细小病毒B19感染真皮成纤维细胞可导致细胞衰老,表现出更高水平的DNA链断裂和氧化损伤,导致衰老相关表型因子的释放,可能在SSc纤维化中发挥作用[41]。
此外,微生物组也是SSc的潜在致病触发或驱动因素。已有研究总结了SSc患者肠道、皮肤和肺部微生物群的改变,包括肠道粪杆菌等共生属数量减少以及梭杆菌等致病菌增加。肠道菌群相关代谢物三甲胺n-氧化物在SSc特别是晚期器官受累的SSc患者中升高,提示微生物稳态失调可能是SSc发生发展的因素之一,相关研究的持续进展将为SSc发病机制的分子途径提供有价值的见解[42-43]。
3.5 干燥综合征
干燥综合征(SS)是一种以泪腺和唾液腺功能下降伴由此导致的眼干和口干为特征的慢性AID。研究显示呼吸系统感染均显著增加了抗SSa和抗SSb抗体阳性SS的发生风险,皮肤和泌尿系统感染与自身抗体阳性的SS发生显著相关;感染次数与自身抗体阳性SS的发生风险呈剂量-反应关系[44]。
迄今为止,EB病毒是SS中研究最多的病原体。EB病毒编码的蛋白可模拟B细胞受体(BCR)和CD40信号通路,刺激相关信号通路,造成SS中B细胞过度激活。在生发中心中,相当比例的滤泡周围浆细胞感染EB病毒显示出对SS自身抗原Ro52的反应性,表明EB病毒感染与自身免疫的持续存在相关联[45]。
此外,EB病毒编码的小RNA可与SS自身抗原La/SSB结合,激活Toll样受体3并导致Ⅰ型干扰素和促炎细胞因子的产生[46]。其他与SS发病风险研究密切相关的病毒还包括CMV、丙型肝炎病毒、柯萨奇病毒和人嗜T淋巴球病毒1型(HTLV-1)[10]。研究显示,肠道生态失调状态也与SS患者较高的疾病活动度相关[47]。
4 新型冠状病毒感染与自身免疫
自从COVID-19疫情暴发以来,大量研究表明COVID-19患者体内存在多种自身抗体,包括抗核抗体、抗磷脂抗体、抗Ro/SSa等[48]。COVID-19患者多种高滴度抗磷脂抗体的出现加重凝血异常,引发类似于抗磷脂综合征(APS)的自身免疫状况,被称为“COVID-19诱导的APS样综合征”[49-50]。
COVID-19的多方面特征与抗黑色素瘤分化相关基因5(MDA5)抗体相关的皮肌炎表现也有相似性,COVID-19患者血浆中存在抗MDA5抗体,抗体阳性患者表现为重症、低生存期、长病程、高器官功能障碍发生率、高龄等,其中抗MDA5抗体滴度≥10.0 U/mL可作为COVID-19重症的强预测因子[51]。
此外,多项研究发现重症COVID-19患者血液中存在1型干扰素自身抗体,可损害下呼吸道1型干扰素免疫并中和其阻断新型冠状病毒感染的能力[52],这与患者的疾病严重度和不良预后有关,因此在确诊COVID-19后应及早进行该自身抗体的检测,从而早期识别危重患者[53]。
新型冠状病毒感染后,部分患者出现AID表现,新发糖尿病风险显著增加[54-55]。新型冠状病毒进入人类宿主细胞不仅结合细胞表面受体血管紧张素转换酶2(ACE2),还需相关进入因子如神经毡蛋白1(NRP1)[56-57]的介导。
研究发现ACE2和NRP1在β细胞中表达,其中NRP1选择性高表达;SARS-CoV-2可感染人胰岛β细胞,降低胰腺胰岛素水平和分泌,并诱导β细胞凋亡,而抑制NRP1可减缓上述进程[58]。
然而,由于短暂的高血糖与新型冠状病毒感染有关,尚不能排除在感染急性期误诊T1DM而造成的假性关联[59]。COVID-19后新发其他AID包括SLE[60]、自身免疫性脑炎[61]、自身免疫溶血性贫血[62]等。
5 研究方法与平台
噬菌体免疫沉淀测序(PhIP-seq)技术是近年来新兴的高通量筛选技术,其联合高容量的噬菌体展示多肽库、抗体免疫沉淀以及高通量测序技术,可进行基于抗原-抗体匹配的、全局性、系统性的抗体库分析。其中,VirScan技术是基于PhIP-seq的一种代表性新型研究平台,可全面无偏倚地检测针对已知感染人类的病毒基因组中编码的所有线性肽抗体,有望广泛用于研究AID中针对病毒抗原的抗体组及特异性结合表位,以高通量、无偏倚的方式筛选与AID有潜在关联的病原微生物及抗原位点。
近年来,基于该技术已发表多篇AID血清抗体组学研究。聚焦于炎症性肠病的研究发现,针对细菌鞭毛素的抗体反应在克罗恩病中占主导地位,且与回肠病变、纤维增生狭窄性疾病及抗酵母菌抗体阳性相关[63]。基于该技术也对MS患者的多种病毒抗体谱(包括EB病毒)进行了检测和病因学探索[21,64]。
6 小 结
虽然研究结果不尽相同,但相关证据为感染与AID致病间的相关性提供了重要支持。值得注意的是,尽管人群中病原微生物感染较为常见,但仅少数人发生AID。除遗传差异可引起机体不同反应外,多种微生物混合感染可能引发自身炎症和自身免疫反应,促进AID的发生发展。
采用靶向一种病原体的单一检测方法确定引起AID的特异性感染病原体极为困难,尤其是当目标病原体流行率较低时,因此需要VirScan等无偏倚的方式寻找AID潜在的相关感染因子。此外,前瞻性研究或小鼠模型的构建,将有助于排除偶然因素,并进一步确定感染与AID致病之间的因果关系。
总之,AID是遗传因素和环境因素相互作用的结果,致病机制非常复杂,在无法改变遗传因素的情况下,通过有效的预防手段减少对感染的暴露,和/或联合应用抗生素可能是降低AID发病率或其严重程度的有效方法。未来需通过更深入的研究以制订合理、高效的治疗策略,提高AID患者的整体生活质量。
参考文献
[1]Pisetsky DS. Pathogenesis of autoimmune disease[J]. Nat Rev Nephrol,2023,19:509-524.
[2]Hussein HM, Rahal EA. The role of viral infections in the development of autoimmune diseases[J]. Crit Rev Microbiol,2019,45:394-412.
[3]Goldberg E, Krause I. Infection and type 1 diabetes mellitus-a two edged sword?[J]. Autoimmun Rev,2009,8:682-686.
[4]Kumagi T, Abe M, Ikeda Y, et al. Infection as a risk factor in the pathogenesis of primary biliary cirrhosis: pros and cons[J]. Dis Markers,2010,29:313-321.
[5]Khalesi Z, Tamrchi V, Razizadeh MH, et al. Association between human herpesviruses and multiple sclerosis: A systematic review and meta-analysis[J]. Microb Pathog,2023,177:106031.
[6]Quaglia M, Merlotti G, De Andrea M, et al. Viral Infections and Systemic Lupus Erythematosus: New Players in an Old Story[J]. Viruses,2021,13:277.
[7]Gremese E, Tolusso B, Bruno D, et al. Infectious agents breaking the immunological tolerance: The holy grail in rheumatoid arthritis reconsidered[J]. Autoimmun Rev,2022,21:103102.
[8]Cheng L, Zhan H, Liu Y, et al. Infectious agents and pathogenesis of Behçet's disease: An extensive review[J]. Clin Immunol,2023,251:109631.
[9]Soldan SS, Lieberman PM. Epstein-Barr virus and multiple sclerosis[J]. Nat Rev Microbiol,2023,21:51-64.
[10]Björk A, Mofors J, Wahren-Herlenius M. Environmental factors in the pathogenesis of primary Sjögren's syndrome[J]. J Intern Med,2020,287:475-492.
[11]Rojas M, Restrepo-Jiménez P, Monsalve DM, et al. Molecular mimicry and autoimmunity[J]. J Autoimmun,2018,95:100-123.
[12]Cunningham MW. Molecular Mimicry, Autoimmunity, and Infection: The Cross-Reactive Antigens of Group A Streptococci and their Sequelae[J]. Microbiol Spectr,2019,7:10.1128/microbiolspec.GPP3-0045-2018.
[13]Lee H, Jeong S, Shin EC. Significance of bystander T cell activation in microbial infection[J]. Nat Immunol,2022,23:13-22.
[14]Cornaby C, Gibbons L, Mayhew V, et al. B cell epitope spreading: mechanisms and contribution to autoimmune diseases[J]. Immunol Lett,2015,163:56-68.
[15]Christen U. Pathogen infection and autoimmune disease[J]. Clin Exp Immunol,2019,195:10-14.
[16]Nekoua MP, Alidjinou EK, Hober D. Persistent coxsackievirus B infection and pathogenesis of type 1 diabetes mellitus[J]. Nat Rev Endocrinol,2022,18:503-516.
[17]Carré A, Vecchio F, Flodström-Tullberg M, et al. Coxsackievirus and Type 1 Diabetes: Diabetogenic Mechanisms and Implications for Prevention[J]. Endocr Rev,2023,44:737-751.
[18]Root-Bernstein R, Chiles K, Huber J, et al. Clostridia and Enteroviruses as Synergistic Triggers of Type 1 Diabetes Mellitus[J]. Int J Mol Sci,2023,24:8336.
[19]Prince MI, Ducker SJ, James OF. Case-control studies of risk factors for primary biliary cirrhosis in two United Kingdom populations[J]. Gut,2010,59:508-512.
[20]Tanaka A, Leung PSC, Gershwin ME. Pathogen infections and primary biliary cholangitis[J]. Clin Exp Immunol,2019,195:25-34.
[21]Bjornevik K, Cortese M, Healy BC, et al. Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis[J]. Science,2022,375:296-301.
[22]Robinson WH, Steinman L. Epstein-Barr virus and multiple sclerosis[J]. Science,2022,375:264-265.
[23]He R, Du Y, Wang C. Epstein-Barr virus infection: the leading cause of multiple sclerosis[J]. Signal Transduct Target Ther,2022,7:239.
[24]Rostgaard K, Nielsen NM, Melbye M, et al. Siblings reduce multiple sclerosis risk by preventing delayed primary Epstein-Barr virus infection[J]. Brain,2023,146:1993-2002.
[25]Afrasiabi A, Keane JT, Ong LTC, et al. Genetic and transcriptomic analyses support a switch to lytic phase in Epstein Barr virus infection as an important driver in developing Systemic Lupus Erythematosus[J]. J Autoimmun,2022,127:102781.
[26]Harley JB, James JA. Epstein-Barr virus infection induces lupus autoimmunity[J]. Bull NYU Hosp Jt Dis,2006,64:45-50.
[27]Reis AD, Mudinutti C, de Freitas Peigo M, et al. Active human herpesvirus infections in adults with systemic lupus erythematosus and correlation with the SLEDAI score[J]. Adv Rheumatol,2020,60:42.
[28]Mahroum N, Elsalti A, Shoenfeld Y. Herpes simplex virus and SLE: Though uncommon yet with significant implications[J]. J Med Virol,2023,95:e28689.
[29]Tomofuji Y, Maeda Y, Oguro-Igashira E, et al. Metagenome-wide association study revealed disease-specific landscape of the gut microbiome of systemic lupus erythematosus in Japanese[J]. Ann Rheum Dis,2021,80:1575-1583.
[30]Aletaha D, Smolen JS. Diagnosis and Management of Rheumatoid Arthritis: A Review[J]. JAMA,2018,320:1360-1372.
[31]Maeda Y, Kurakawa T, Umemoto E, et al. Dysbiosis Contributes to Arthritis Development via Activation of Autoreac-tive T Cells in the Intestine[J]. Arthritis Rheumatol,2016,68:2646-2661.
[32]Wegner N, Wait R, Sroka A, et al. Peptidylarginine deiminase from Porphyromonas gingivalis citrullinates human fibrinogen and α-enolase: implications for autoimmunity in rheumatoid arthritis[J]. Arthritis Rheum,2010,62:2662-2672.
[33]Jiang L, Shang M, Yu S, et al. A high-fiber diet synergizes with Prevotella copri and exacerbates rheumatoid arthritis[J]. Cell Mol Immunol,2022,19:1414-1424.
[34]Zheng Z, Sohn S, Ahn KJ, et al. Serum reactivity against herpes simplex virus type 1 UL48 protein in Behçet's disease patients and a Behçet's disease-like mouse model[J]. Acta Derm Venereol,2015,95:952-958.
[35]Silva NSM, Rodrigues LFC, Dores-Silva PR, et al. Structural, thermodynamic and functional studies of human 71 kDa heat shock cognate protein (HSPA8/hHsc70)[J]. Biochim Biophys Acta Proteins Proteom,2021,1869:140719.
[36]Yang TH, Aosai F, Norose K, et al. Heat shock cognate protein 71-associated peptides function as an epitope for Toxoplasma gondii-specific CD4+CTL[J]. Microbiol Immunol,1997,41:553-561.
[37]Cho SB, Zheng Z, Ahn KJ, et al. Serum IgA reactivity against GroEL of Streptococcus sanguinis and human heterogeneous nuclear ribonucleoprotein A2/B1 in patients with Behçet disease[J]. Br J Dermatol,2013,168:977-983.
[38]Deniz R, Emrence Z, Yalçinkaya Y, et al. Improved sensitivity of the skin pathergy test with polysaccharide pneumococcal vaccine antigens in the diagnosis of Behçet disease[J]. Rheumatology (Oxford),2023,62:1903-1909.
[39]Ouchene L, Muntyanu A, Lavoué J, et al. Toward Understanding of Environmental Risk Factors in Systemic Sclerosis[J]. J Cutan Med Surg,2021,25:188-204.
[40]Soffritti I, D'Accolti M, Maccari C, et al. Human Cytomegalovirus and Human Herpesvirus 6 Coinfection of Dermal Fibroblasts Enhances the Pro-Inflammatory Pathway Predisposing to Fibrosis: The Possible Impact on Systemic Sclerosis[J]. Microorganisms,2022,10:1600.
[41]Arvia R, Zakrzewska K, Giovannelli L, et al. Parvovirus B19 induces cellular senescence in human dermal fibroblasts: putative role in systemic sclerosis-associated fibrosis[J]. Rheumatology (Oxford),2022,61:3864-3874.
[42]Kim S, Park HJ, Lee SI. The Microbiome in Systemic Sclerosis: Pathophysiology and Therapeutic Potential[J]. Int J Mol Sci,2022,23:16154.
[43]Stec A, Maciejewska M, Paralusz-Stec K, et al. The Gut Microbial Metabolite Trimethylamine N-Oxide is Linked to Specific Complications of Systemic Sclerosis[J]. J Inflamm Res,2023,16:1895-1904.
[44]Mofors J, Arkema EV, Björk A, et al. Infections increase the risk of developing Sjögren's syndrome[J]. J Intern Med,2019,285:670-680.
[45]Croia C, Astorri E, Murray-Brown W, et al. Implication of Epstein-Barr virus infection in disease-specific autoreactive B cell activation in ectopic lymphoid structures of Sjögren's syndrome[J]. Arthritis Rheumatol,2014,66:2545-2557.
[46]Iwakiri D, Zhou L, Samanta M, et al. Epstein-Barr virus (EBV)-encoded small RNA is released from EBV-infected cells and activates signaling from Toll-like receptor 3[J]. J Exp Med,2009,206:2091-2099.
[47]de Paiva CS, Jones DB, Stern ME, et al. Altered Mucosal Microbiome Diversity and Disease Severity in Sjögren Syndrome[J]. Sci Rep,2016,6:23561.
[48]Liu Y, Sawalha AH, Lu Q. COVID-19 and autoimmune diseases[J]. Curr Opin Rheumatol,2021,33:155-162.
[49]Zhang Y, Xiao M, Zhang S, et al. Coagulopathy and Antiphospholipid Antibodies in Patients with COVID-19[J]. N Engl J Med,2020,382:e38.
[50]Xiao M, Zhang Y, Zhang S, et al. Antiphospholipid Antibodies in Critically Ill Patients With COVID-19[J]. Arthritis Rheumatol,2020,72:1998-2004.
[51]Wang G, Wang Q, Wang Y, et al. Presence of Anti-MDA5 Antibody and Its Value for the Clinical Assessment in Patients With COVID-19: A Retrospective Cohort Study[J]. Front Immunol,2021,12:791348.
[52]Philippot Q, Fekkar A, Gervais A, et al. Autoantibodies Neutralizing Type I IFNs in the Bronchoalveolar Lavage of at Least 10% of Patients During Life-Threatening COVID-19 Pneumonia[J]. J Clin Immunol,2023,43:1093-1103.
[53]Solanich X, Rigo-Bonnin R, Gumucio VD, et al. Pre-existing Autoantibodies Neutralizing High Concentrations of Type I Interferons in Almost 10% of COVID-19 Patients Admitted to Intensive Care in Barcelona[J]. J Clin Immunol,2021,41:1733-1744.
[54]Barrett CE, Koyama AK, Alvarez P, et al. Risk for Newly Diagnosed Diabetes >30 Days After SARS-CoV-2 Infection Among Persons Aged <18 Years- United States, March 1, 2020-June 28, 2021[J]. MMWR Morb Mortal Wkly Rep,2022,71:59-65.
[55]McKeigue PM, McGurnaghan S, Blackbourn L, et al. Relation of Incident Type 1 Diabetes to Recent COVID-19 Infection: Cohort Study Using e-Health Record Linkage in Scotland[J]. Diabetes Care,2023,46:921-928.
[56]Daly JL, Simonetti B, Klein K, et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection[J]. Science,2020,370:861-865.
[57]Cantuti-Castelvetri L, Ojha R, Pedro LD, et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity[J]. Science,2020,370:856-860.
[58]Wu CT, Lidsky PV, Xiao Y, et al. SARS-CoV-2 infects human pancreatic β cells and elicits β cell impairment[J]. Cell Metab,2021,33:1565-1576.e5.
[59]Chen J, Wu C, Wang X, et al. The Impact of COVID-19 on Blood Glucose: A Systematic Review and Meta-Analysis[J]. Front Endocrinol (Lausanne),2020,11:574541.
[60]Bonometti R, Sacchi MC, Stobbione P, et al. The first case of systemic lupus erythematosus (SLE) triggered by COVID-19 infection[J]. Eur Rev Med Pharmacol Sci,2020,24:9695-9697.
[61]Valencia Sanchez C, Theel E, Binnicker M, et al. Autoimmune Encephalitis After SARS-CoV-2 Infection: Case Frequency, Findings, and Outcomes[J]. Neurology,2021,97:e2262-e2268.
[62]Capes A, Bailly S, Hantson P, et al. COVID-19 infection associated with autoimmune hemolytic anemia[J]. Ann Hematol,2020,99:1679-1680.
[63]Bourgonje AR, Andreu-Sánchez S, Vogl T, et al. Phage-display immunoprecipitation sequencing of the antibody epitope repertoire in inflammatory bowel disease reveals distinct antibody signatures[J]. Immunity,2023,56:1393-1409.e6.
[64]Enose-Akahata Y, Wang L, Almsned F, et al. The repertoire of CSF antiviral antibodies in patients with neuroinflammatory diseases[J]. Sci Adv,2023,9:eabq6978.
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言